This article has since been published in the University of Sheffield’s Biology magazine. That version can be accessed at the end of this article.
“Over the oxygen supply of the body carbon dioxide spreads its protecting wings.” – Friedrich Miescher, Swiss physician, 1885
Carbon dioxide is usually seen only as a waste gas, while lactate is often treated as a simple by-product of exertion during exercise, and nothing more. In reality, the balance between the two may reveal something deeper about whether metabolism is efficient, oxidative, and protective, or stressed, inflammatory, and degenerative.
CO2 vs Lactate
Ordinarily, carbon dioxide (and bicarbonate) is typically thought of with respect to the regulation of pH, and only in a very general sense, but its essential and protective roles extend far beyond this. Because carbon dioxide plays an important role in keeping the pH of the blood within a narrow range, it has been commonly thought of as a toxin, since a supraphysiological excess can cause unconsciousness and acidosis. Increasing carbon dioxide, however, doesn’t necessarily cause acidosis, and acidosis caused by carbon dioxide isn’t nearly as harmful as lactic acidosis – the accumulation of too much lactic acid in the body, often due to a lack of carbon dioxide (Kamel et al., 2019).
Carbon dioxide and lactate appear to have an inverse relationship, most likely explained by the “Randle cycle”. In 1963, the British physiologist Sir Philip Randle, researching at Oxford University at the time, observed that glucose and fatty acids compete for oxidation in the cell (Randle et al., 1963). He realised that the mitochondrion can only oxidise either glucose or fatty acids at any given time, not both. Therefore, if an influx of fatty acids are present in the bloodstream, whether that be due to stress, inflammation, or just dietary intake, glucose gets “outcompeted” for oxidation in the mitochondria. As a result, much of the remaining glucose can only be burnt through glycolysis, a less efficient process which occurs outside of the mitochondria, rather than through efficient oxidative phosphorylation in the mitochondria. Glycolysis produces no carbon dioxide, rather producing lactate instead. The complete metabolism of glucose, critically involving oxidative phosphorylation, produces more carbon dioxide (and no lactate) per molecule of oxygen compared to glycolysis, or the oxidation of fatty acids.
This is important. This is because carbon dioxide is protective, since it begets effective metabolism, and thus better structure and function and so on. More carbon dioxide in our blood leads to a more efficient usage of oxygen, through the Bohr effect, and an improved blood supply to the tissues due to its function as a vasodilator.
A reduction of carbon dioxide however, caused by hyperventilation for example, can provoke seizure, cause muscle spasms (due to an inability to relax the muscle, which is a high-energy requiring state), promote vascular leakiness, and contribute towards inflammation and potentially even thrombosis events (Salvati and Beenhakker, 2019; Lewis, 2025; Nagy et al., 1985, Curley et al., 2010; Dai et al., 2006; Staubli et al., 1994). Small fluctuations in serum oxygen and carbon dioxide levels occur many times throughout the day, but chronically reduced carbon dioxide – or chronically elevated lactate – can have more serious consequences.
Acute lactate rises (like from exercise) aren’t necessarily concerning, but chronically elevated lactate seen in hypoxia and poor metabolic states leads to pro-inflammatory signalling, cell proliferation (and thus tumour growth), fibrosis, and cardiolipin disruption (Kawauchi, et al., 2008; Liu et al., 2023; Longhitano et al., 2022; Sun et al., 2024; San-Millán et al., 2022). Cardiolipin is a unique phospholipid and is required for the optimal activity of inner mitochondrial membrane proteins, including the enzyme complexes of the electron transport chain and ATP production (Paradies et al., 2019). Cardiolipin also plays a role in mitochondrial membrane morphology, stability, dynamics, and in mitochondrial biogenesis (which primarily occurs during sleep when CO2 levels rise). Therefore, it is no surprise that the effects of chronically elevated lactate levels on mitochondrial function may be why it is so strongly associated with type II diabetes (Crawford et al., 2010).
Otto Warburg won the Nobel Prize in Physiology in 1931 for his discovery of the “Warburg Effect”, referring to the observation that many cancer cells produce large quantities of lactate, with glucose not being completely oxidised (into carbon dioxide; Warburg, 1956). In the cases of these tumours, given that they are predominantly running on glycolysis – not the efficient oxidative phosphorylation process – focusing on restoring effective respiration would be the most sensible step in addressing Warburg Effect-displaying tumours, as well as similar diseases involving degenerative metabolism and inflammation.
Examples in nature
Bats have a very high metabolism, as well as an extremely long lifespan for an animal of their size and weight. While similarly-sized animals tend to live for only a few years, bats can often live for a few decades. One plausible explanation for this has to do with their caves. Bat caves typically have more carbon dioxide than the outside atmosphere, but they also usually contain a large amount of ammonia. However, bats maintain a high level of carbon dioxide in their blood, which protects them from the otherwise toxic effects of the ammonia (ammonia also promotes glycolysis and lactate production). Given that higher altitude is strongly positively correlated with lower mortality (Burtscher, 2014), and so is carbon dioxide saturation with altitude, it wouldn’t be much of a stretch to say that carbon dioxide is having a protective effect on bat lifespan.
Similarly, naked mole rats are another small animal with a very long lifespan. In captivity they live up to 30 years old, and a little shorter in the wild, roughly 9 times longer than similarly-sized mice. They also live underground, in closed burrows with higher carbon dioxide levels. Native to the high-altitude plains of Ethiopia and Kenya, they would naturally have higher carbon dioxide levels in their tissues, perhaps helping explain their long lifespans.
Like mole rats, bees also live in closed, high carbon dioxide environments. Interestingly, in colonies of either organism, there is a queen, and it is her who is the only female that reproduces. The queen bee is the largest individual in the colony, and her workers happen to carefully control the concentration of carbon dioxide, because it must be kept high in order to accelerate ovary action and induce egg laying (Koywiwattrakul et al., 2005). Since queen bees and mole rats live in the dark, their high carbon dioxide levels probably compensate for the lack of light, because both light and CO2 help to maintain oxidative metabolism and inhibit lactate formation.
In humans, it has been observed that higher carbon dioxide levels, at altitude, can protect against obesity and metabolic complications. Obese adults with metabolic syndrome – who could eat freely and were not allowed to exercise – were exposed to an altitude of 2,650 m, and their metabolic rates increased by an average of about 20% and roughly 3 kgs were lost by each subject after only 2 weeks (Lipp et al., 2010). Similarly, Butterfield et al. (1992) observed that adults taken to 4,300 m showed a 27% rise in basal metabolic rate on only the second day of testing. On average, the subjects ate 452 calories more than usual per day, yet still lost over 2.1 kg of adipose tissue after 2 weeks. Interestingly, during strenuous exercise, the subjects were able to expend a whopping 37% more energy than at sea level (Butterfield et al., 1992). This likely explains why elite endurance athletes tend to train at altitude before racing season.
Furthermore, construction workers in India who transferred from low altitude areas to working at 2,750 m in the Himalayas maintained their body weight despite eating between 400-2,900 extra calories per day (Sridharan et al. , 1987).
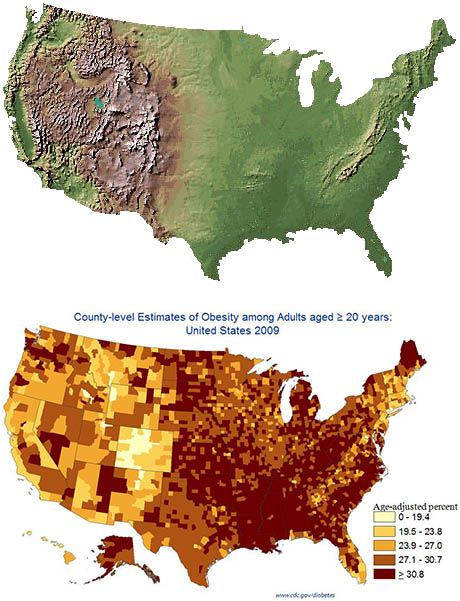
A 2025 systematic review and meta-analysis concluded that moderate passive hypoxic conditioning, in other words, increasing one’s tolerance to carbon dioxide, could be an effective therapy in managing adipose mass without potential health risks, like those seen in other current therapies (Timon et al., 2025). Figure 1 compares the topography of the United States with geographically distributed obesity rates. Coincidence?
While altitude itself is a complex stimulus and not necessarily equivalent to carbon dioxide therapy, these findings are still consistent with the broader idea that metabolism can be shifted away from the low-energy, lactate-dominant state typical of obesity and related degenerative conditions and towards a more efficient, oxidative state.
Figure 1. Composite illustration used in blog coverage of Voss et al. (2013), combining a U.S. topographic relief map (USGS National Elevation Dataset) with the CDC’s 2009 county-level age-adjusted adult obesity map.
Conclusion
Taken together, it is clear to see that carbon dioxide’s functions stretch far beyond blood acidity, and I believe it shouldn’t be merely viewed as a waste product of mitochondrial respiration. Carbon dioxide appears to have an evolutionary alignment with our physiology, as well as with those in other organisms, and its numerous actions on fundamental biological processes in a positive and coherent way is what constitutes its broad protective role. Restoring the conditions that allow carbon dioxide to be produced and retained may likely prove to be a fundamental and safe way we can restore metabolic health and protect ourselves from degenerative diseases.
Butterfield, G.E., Gates, J., Fleming, S., Brooks, G.A., Sutton, J.R. and Reeves, J.T. (1992). Increased energy intake minimizes weight loss in men at high altitude. Journal of Applied Physiology, 72(5), pp.1741–1748. doi:https://doi.org/10.1152/jappl.1992.72.5.1741
Curley, G., Laffey, J.G. and Kavanagh, B.P. (2010). Bench-to-bedside review: carbon dioxide. Critical care (London, England), [online] 14(2), p.220. doi:https://doi.org/10.1186/cc8926
Crawford, S.O., Hoogeveen, R.C., Brancati, F.L., Astor, B.C., Ballantyne, C.M., Schmidt, M.I. and Young, J.H. (2010). Association of blood lactate with type 2 diabetes: the Atherosclerosis Risk in Communities Carotid MRI Study. International Journal of Epidemiology, [online] 39(6), pp.1647–1655. doi:https://doi.org/10.1093/ije/dyq126
Kamel, K.S., Oh, M.S. and Halperin, M.L. (2019). L-lactic acidosis: pathophysiology, classification, and causes; emphasis on biochemical and metabolic basis. Kidney International, 97(1). doi:https://doi.org/10.1016/j.kint.2019.08.023
Koywiwattrakul, P., Thompson, G.J., Sitthipraneed, S., Oldroyd, B.P. and Maleszka, R. (2005). Effects of carbon dioxide narcosis on ovary activation and gene expression in worker honeybees, Apis mellifera. Journal of Insect Science, 5(1). doi:https://doi.org/10.1093/jis/5.1.36
Lippl, F.J., Neubauer, S., Schipfer, S., Lichter, N., Tufman, A., Otto, B. and Fischer, R. (2010). Hypobaric Hypoxia Causes Body Weight Reduction in Obese Subjects. Obesity, 18(4), pp.675–681. doi:https://doi.org/10.1038/oby.2009.509
Liu, W., Wang, Y., Bozi, L.H.M., Fischer, P., Jedrychowski, M.P., Xiao, H., Wu, T., Darabedian, N., He, X., Mills, E.L., Burger, N., Shin, S., Reddy, A., Sprenger, H.-G., Tran, N., Winther, S., Hinshaw, S.M., Shen, J., Seo, H.-S. and Song, K. (2023). Lactate regulates cell cycle by remodeling the anaphase promoting complex. Nature. [online] doi:https://doi.org/10.1038/s41586-023-05939-3
Longhitano, L., Vicario, N., Tibullo, D., Cesarina Giallongo, Broggi, G., Caltabiano, R., Maria, G., Altieri, R., Baghini, M., Michelino Di Rosa, Parenti, R., Giordano, A., Maria Caterina Mione and Giovanni Li Volti (2022). Lactate Induces the Expressions of MCT1 and HCAR1 to Promote Tumor Growth and Progression in Glioblastoma. Frontiers in Oncology, 12. doi:https://doi.org/10.3389/fonc.2022.871798
Paradies, G., Paradies, V., Ruggiero, F.M. and Petrosillo, G. (2019). Role of Cardiolipin in Mitochondrial Function and Dynamics in Health and Disease: Molecular and Pharmacological Aspects. Cells, 8(7), p.728. doi:https://doi.org/10.3390/cells8070728
Randle, P.J., Garland, P.B., Hales, C.N. and Newsholme, E.A. (1963). THE GLUCOSE FATTY-ACID CYCLE ITS ROLE IN INSULIN SENSITIVITY AND THE METABOLIC DISTURBANCES OF DIABETES MELLITUS. The Lancet, 281(7285), pp.785–789. doi:https://doi.org/10.1016/s0140-6736(63)91500-9
Salvati, K.A. and Beenhakker, M.P. (2019). Out of thin air: Hyperventilation-triggered seizures. Brain research, [online] 1703, pp.41–52. doi:https://doi.org/10.1016/j.brainres.2017.12.037
Sridharan, K., A.K. Mukherjee, S.K. Grover, M.M.L. Kumaria, B.S. Arora, and R.M. Rai 1987. Assessment of nutritional status and physical work capacity of road construction workers at altitude of 2,150–2,750 m on two different ration scales. Nutr. Rep. Int. 35:1269–1277
Iñigo San-Millán, Sparagna, G.C., Chapman, H.L., Warkins, V.L., Chatfield, K.C., Shuff, S., Martinez, J.L. and Brooks, G.A. (2022). Chronic Lactate Exposure Decreases Mitochondrial Function by Inhibition of Fatty Acid Uptake and Cardiolipin Alterations in Neonatal Rat Cardiomyocytes. Frontiers in Nutrition, [online] 9. doi:https://doi.org/10.3389/fnut.2022.809485
Staubli, M., Vogel, F., Bartsch, P., Fluckiger, G. and Ziegler, W.H. (1994). Hyperventilation-induced changes of blood cell counts depend on hypocapnia. European Journal of Applied Physiology and Occupational Physiology, 69(5), pp.402–407. doi:https://doi.org/10.1007/bf00865403
Sun, Z., Ji, Z., Meng, H., He, W., Li, B., Pan, X., Zhou, Y. and Yu, G. (2024). Lactate facilitated mitochondrial fission-derived ROS to promote pulmonary fibrosis via ERK/DRP-1 signaling. Journal of Translational Medicine, [online] 22, p.479. doi:https://doi.org/10.1186/s12967-024-05289-2
Timon, R., Rojas-Valverde, D. and Camacho-Cardenosa, M. (2025). Effect of passive hypoxia on body fat mass in older and young adults: A systematic review and meta-analysis. Physiology & Behavior, pp.114948–114948. doi:https://doi.org/10.1016/j.physbeh.2025.114948
Voss JD, Masuoka P, Webber BJ, Scher AI, Atkinson RL. Association of elevation, urbanization and ambient temperature with obesity prevalence in the United States. International Journal of Obesity. 2013;37(10):1407–1412
This article has since been published in The War Kitchen Magazine (Issue 41). This version can be viewed at the end of this article.
What is endotoxin?
It is probably quite likely that the reader has never heard of endotoxin before. Endotoxin, specifically lipopolysaccharide (LPS), the most potent form, is a toxic component found in the outer membrane of Gram-negative bacteria. When these “bad” bacteria die, or are destroyed in our guts by white blood cells, these endotoxins break off and can enter the circulating bloodstream (a process known as endotoxemia). LPS has a very high binding affinity for the TLR4 receptor, which in turn creates inflammation, and has been implicated in many chronic diseases.
The harmful effects of endotoxin on energy and structure
Endotoxin is, unfortunately, somewhat uniquely evil in how many biological systems it can impair. The main toxic mechanism of LPS involves inappropriate excitation of cells, followed by inflammation and mitochondrial dysfunction. This inflammation causes cells to swell, taking up water and losing their structure. When a cell lacks structure, it also lacks energy, and vice versa. Think of a tidy home; that order (structure) took energy. When cells lack order, they lack the means and infrastructure to generate energy, gradually resulting in less and less structure and energy. High-energy cells are highly specialised and differentiated, and it’s that abundance of cellular energy which allowed them to develop and carry out their specialised roles in the first place. Therefore, when mitochondrial function is impaired by LPS (whether through cytokine release, accumulation of excess oestrogen, or the swelling-induced mechanical distortion of mitochondria) and cellular energy is decreased, cells can no longer carry out their individual roles and the whole organism suffers, leading to disease. These faulty, impaired mitochondria produce more reactive oxygen species (ROS), which in turn creates more DNA damage, eventually potentially leading to cancer.
Hummingbirds have the highest metabolic rate per gram of body weight of any animal, roughly 50 times higher than an elephant and 10 times higher than a pigeon. Their beautiful colours and patterns reflect their highly differentiated nature and it’s their powerful metabolism, and thus abundance of cellular energy, which has allowed them to develop these beautiful structures.
Endotoxin and Cancer: Why we shouldn’t overlook it
“I’m inclined to think that even cancer is largely an endotoxin problem. Chronic, year after year, bad food, irritating the intestine, causing increased histamine and serotonin and oestrogen. A constant stream of endotoxin flowing through the system, shaking up chromosomes, creating inflammation, fatiguing the repair systems. Finally, when the repair systems are down, to a certain extent, the cancers that are constantly popping up stop being removed. And so if you don’t die of acute sepsis, then heart disease and cancer are the result of chronic exposure to the endotoxin.” – Dr Raymond Peat
Until only fairly recently, endotoxin was not suspected to be involved in cancer. Fortunately, now we know that not only can it provide a favourable environment for cancers to thrive in, but it can even directly cause tumours to form.
One of the ways in which chronic endotoxin exposure can lead to cancer formation is through its ability to allow oestrogen to accumulate in excess, and thus skew its relationship with the opposing factors like thyroid hormone and progesterone. Endotoxin’s inhibition of the glucuronidation system (the system which helps export substances from the body through urine or faeces via bile) leads to oestrogen accumulation in our cells, as well as prolonging circulation of endotoxin in the body (Bánhegyi, et al., 1995). The liver is constantly inactivating hormones, like oestrogen, and when this ability is shut down, it can no longer stabilise hormone balances as normal. Excess oestrogen (also known as ‘oestrogen dominance’) has been shown to play roles in breast, ovarian, prostate, uterine, pituitary, lung, and skin cancers (Steinberg et al., 1994; Langdon et al., 2020; Lafront et al., 2024; Grady et al., 1995; Heaney, Fernando and Melmed, 2002; Li, Pei and Feng, 2023; Smida et al., 2020).
Excess oestrogen can be metabolised into catechol oestrogens which mimic compounds such as adrenaline, and it’s in these forms when they can be oxidised into quinones which produce toxic free radicals, leading to tumour formation (Cavalieri and Rogan, 2021).
Furthermore, LPS forces macrophages and the Kupffer cells in the liver to synthesise more tumour necrosis factor (TNF). Good, right? Unfortunately not. Despite its name, in reality TNF significantly enhances tumour growth and metastasis of numerous cancers (Wang and Lin, 2008). I think a more appropriate name is needed.
It’s important and helpful to look at endotoxin as this chronic, baseline stressor on the body. Like the quote above suggests, when your gut lining is damaged, you have a constant flow of endotoxin being absorbed and creating inflammation. At these sites of inflammation, immune cells become activated, and their evolutionary advantage is that they produce free radicals and inflammatory enzymes to kill pathogens. However, these compounds also break DNA strands. And so when you have chronic inflammation over the long term, although we have DNA repair systems to fix these damages, these systems become overloaded, and mutations accumulate. Have enough of the correct mutations in the right places, and cancers start forming.
Why endotoxin is becoming such a large problem today
Endotoxin and its mechanisms of damage have never been more of a serious issue than they are right now. We consume too much ultra-processed ‘food’ and lace it with biological toxins. Our fields, waters, and skies are riddled with environmental poisons which leach into the animals and crops we eat. And as it turns out, consumption of these ultra-processed foods, emulsifiers, pesticides, and pollutants wages war on our biology. Overconsumption of, and overexposure to these things, unhealthy farming practices, and more generally “stressful modern conditions”, tear apart our epithelial gut lining, increasing permeability and leading to endotoxemia, systemic inflammation, and metabolic disorders (Camilleri et al., 2019; Poto et al., 2023; Do, 2024; Mallardi et al., 2025).
Consequently, young people – who have notoriously bad diets – are becoming ill at higher rates than ever before. And if you have a population wherein both the old and the young are sick, it creates an unsustainable dual strain on the healthcare services and medical resources. In combination, a healthcare system which doesn’t effectively heal the patient but instead suppresses the symptoms of a complicated underlying problem which is never fixed, only ever prolongs the patients’ poor health and thus the financial burden on the state.
One particularly striking study was carried out last year and it explored endotoxin’s impact on human emotions. In the study, the researchers discovered that in the test subjects that were exposed to LPS, there was a significant reduction in people’s empathy for others’ psychological pain (Flasbeck et al., 2024). It was the “endotoxin-induced acute inflammation” which led to the volunteers becoming more callous and less empathetic towards others. Evolutionarily, this makes sense; it likely reflects an adaptive strategy to conserve one’s energy through not responding empathetically when sick oneself. However, it begs an interesting question, that perhaps those who seem to be ‘more apathetic by nature’ are actually just chronically inflamed on the inside?
Endotoxin has also been shown to:
Drive obesity, diabetes, and cardiovascular disease (Perng et al., 2022; Lin et al., 2022)
Accelerate the cognitive decline of ageing (Fei et al., 2022)
Cause chronic inflammation (Shan et al., 2020)
Cause infertility and lower testosterone levels in men (Tremellen, 2016)
Cause infertility and lower progesterone levels in women (Tremellen et al., 2014)
Cause depression and anxiety (Radjabzadeh et al., 2022)
Trigger social withdrawal and isolation (Stockmaier et al., 2020)
Potentially cause cancerisation of normal cells (Tang et al., 2021)
Increase serum oestrogen and cortisol levels (Christeff et al., 2002; Grigoleit et al., 2011)
Accelerate atherosclerosis (Reifenberg et al., 2009; Mastrangelo et al., 2025)
Cause metastasis of some cancers (Cao et al., 2023)
Increase serotonin release from platelets, suppressing mitochondria, and impairing circulation (J. Hawiger et al., 1977)
Attenuate empathy for others (Flasbeck et al., 2024)
Cause muscle weakness/damage (myopathy) (Hashida, Steelman and Erdman, 2024)
Drive Alzheimer’s disease (Pogue et al., 2022; Brown and Heneka, 2024)
… to name but a few. So, it’s hopefully pretty clear that endotoxin is really rather awful for us. But, if it’s so bad, why haven’t I ever heard about it before? Well, I believe it’s largely because our culture and education systems haven’t assimilated many of the scientific discoveries of the past century or so. Endotoxin has been known to be toxic for decades now, but money drives research, and sometimes, if research isn’t deemed to be profitable for investors, said investors are less likely to cough up funding.
It’s not all doom and gloom though, because in light of the heightened interest in microbiome research over the last few decades, researchers are now becoming increasingly curious about endotoxin. Partly as a result, it is now known that there are many simple and effective ways to protect oneself against endotoxin.
Carrots?
Yes, really. Allow me to explain. While the discovery of gut-derived endotoxin as a driver of chronic inflammation has led to the development of pharmaceutical strategies, some have long suggested simple dietary interventions to mitigate their damage. Decades before mainstream science linked endotoxin to chronic disease, biologist and author Dr Raymond Peat suggested that raw carrot fibre could prevent this toxicity through binding and removing endotoxin from the intestine.
“Endotoxin formed in the bowel can block respiration and cause hormone imbalances contributing to instability of the nerves, so it is helpful to optimize bowel flora, for example with a carrot salad; a dressing of vinegar, coconut oil and olive oil, carried into the intestine by the carrot fiber, suppresses bacterial growth while stimulating healing of the wall of the intestine [thus preventing endotoxemia].” – Dr Raymond Peat, PhD
How does it work?
Carrot fibre, specifically raw carrot fibre, acts like a non-digestible, hydrophobic sponge that collects endotoxin through various mechanisms, and escorts it out of the gut before it can be absorbed into the bloodstream. In other words, it acts as a vessel for all sorts of toxins to be cleared from the digestive system.
The insoluble nature of raw carrot possesses hydrophobic regions, which are excellent at interacting with the lipid-rich region of endotoxin, as well as a rigid and porous matrix which is effective at physically trapping LPS. It’s also poorly fermented by gram negative gut bacteria, which means that it doesn’t feed excess bacterial growth, and thus create more endotoxin. Raw fibre is also more resistant to being broken down, therefore it maintains its structural integrity throughout the whole digestive tract, continuing to successfully adsorb endotoxin.
Coconut oil reduces the source of endotoxin by inhibiting gut bacterial overgrowth, and apple cider vinegar also discourages bacterial proliferation and supports digestive efficiency, through lowering the pH of the salad and improving upper GI digestion.
Recipe: The Ray Peat Carrot Salad (Serves one)
1 large raw carrot, peeled lengthwise (or grated)
1 tablespoon apple cider vinegar
1 tablespoon coconut oil (or olive oil, or half of both)
Pinch of salt
What else can be utilised to prevent absorption of endotoxin?
White button mushrooms
Well-cooked, white button mushrooms help to clear endotoxin from the gut because they contain indigestible polysaccharides and beta-glucans that bind LPS in the gut and modulate immune cell activity to reduce inflammation.
It is paramount, though, that the mushrooms are boiled for 1-3 hours (or microwaved for 4-6 minutes). This is because raw or lightly cooked mushrooms contain fungal toxins (e.g. hydrazine) and irritants (e.g. chitin) that can irritate the gut lining and trigger the immune system’s inflammatory cascade. Extended cooking denatures or evaporates away these toxins and that the chitin matrix is softened, reducing the potential for irritation. White button mushrooms are chosen for their very low polyunsaturated fatty acid (PUFA) content, lowering the potential for mitochondrial oxidative stress. You can follow this link for a recipe of mushroom soup, which adheres to the above information.
Coffee
Coffee has numerous protective effects on the gut barrier and can reduce circulating endotoxin levels, regardless of the caffeine concentration. Coffee’s polyphenols and diterpenes strengthen gut lining integrity and, by increasing peristalsis, coffee can reduce constipation-related endotoxemia.
If you want to get really particular about your coffee and extract as much of the ‘good stuff’as possible, such as the terpenoids, trigonelline, melanoidins, chlorogenic acid, and tocopherols etc., you could brew your coffee with ascending heat. This is because these unique beneficial compounds are most soluble at lower temperatures. So, if you dump boiling hot water over the grounds, you’re going to denature a lot of them. Therefore, you could moisten the grounds slightly with warm water first, then slowly add water of increasing temperature, allowing things with different solubilities to periodically release into the water. This process often gives the coffee this deep orange colour and introduces pleasant new aromas.
Orange juice
Fresh, unpasteurised orange juice has been shown to be extremely protective against endotoxin. The vitamin C, flavonoids, and sugars in orange juice all enhance tight junction stability and reduce oxidative stress in the gut. The sugar protects against stress-induced intestinal permeability through preventing cortisol-driven breakdown of tight junctions and supporting liver detoxification of LPS through glycogen replenishment.Naringenin, found in other citrus fruits as well as oranges, is a TLR4 antagonist, meaning it binds to the receptor in a way which prevents the correct substrate from binding, inhibiting the inflammatory process.
A classic study in 2010 discovered that drinking a glass of fresh orange juice alongside a high-fat, high-carbohydrate breakfast meal (“similar to a McDonald’s-style fast food breakfast sandwich”) neutralised the typical proinflammatory response. The control group showed significant postprandial increases in plasma endotoxin and TLR4 expression, but the orange juice group did not (Ghanim et al., 2010).
Glycine
Glycine, an anti-inflammatory amino acid deficient in both modern diets and endogenous production in adults today (Meléndez-Hevia et al., 2009), decreases mortality in animals dosed with endotoxin. Glycine is abundant in gelatinous cuts of meat (think “eating nose to tail”) and plays key roles in maintaining integrity of the gut barrier. Unfortunately, glycine is often replaced by glyphosate (a toxic herbicide) in the epidermis, by mimicking glycine’s structure. It’s this replacement which leads to endotoxemia for many people.
To combine the positive effects of both orange juice and glycine, you could make orange juice gummies with gelatine (being 33% glycine). 600ml orange juice and 5 tablespoons of gelatine is a good place to start. Bloom your gelatine (add slowly) into your orange juice and heat very gently in a saucepan. Transfer to a container once the gelatine is dissolved and cool in the fridge for a couple of hours to set. Add more gelatine for a firmer texture.
Avoid polyunsaturated fatty acids
Polyunsaturated fatty acids (PUFAs; the reason why seed oils are bad for us) are highly prone to lipid peroxidation, and it’s this peroxidation in our gut which damages junctions and creates a ‘leaky gut’, allowing endotoxin to pass into our bloodstream. PUFAs are also precursors to pro-inflammatory mediators like prostaglandins, which intensify the inflammatory cytokine response when endotoxin activates TLR4. It’s likely because of this that septic shock (when the body releases too many cytokines into the bloodstream), caused by the injection of endotoxin, is 100% lethal to animals fed diets high in PUFA, whereas it’s only 24% lethal to PUFA-deficient animals (Cook et al., 1981).
The problem with reductionism in science
Problems like endotoxemia are complicated, and it’s arguably largely for this complexity as well as reductionist thinking in our scientific culture, that it’s taken so long for us to figure it out.
Reductionism, which reduces the explanation for something to a description of its parts, the ways the parts interact, and a small number of physical laws, avoids bothering with physiological complexity in understanding something biological. It’s a way of drawing hypotheses based on a small set of binary principles and therefore ‘proving it scientifically’. With this reductionist mystique of imposing conclusions about things, of claiming that because we already have the knowledge of the parts we can sufficiently explain the problem as a whole, we miss out on the ‘unexpected’ links in disease. We would likely never have connected endotoxin to Alzheimer’s, or atherosclerosis, or cancer with this philosophy. Therefore, as Dr Raymond Peat wrote, “satisfactory solutions to complex problems will be possible only if we directly confront the complexity” (Peat, 2001).
In order to progress scientifically in a qualitative manner, we have to unsubscribe from the reductionist notion of ‘logical deduction’ assigning general theory to solve specific problems, because in reality, biology doesn’t often work like that. We must try to avoid the mindset that nothing else is to be found within problems that have already been ‘solved’ so as to not thwart the development of new branches and ideas in science. This mirrors one of the current problems with academia, particularly in biosciences – that the emphasis is on imprinting information onto the students’ minds to be regurgitated when recalled upon, rather than on applying their own judgement upon the world to cause a material change. Niels Bohr was given the Nobel Prize for his incorrect electron shell model. The important takeaway is not that the model was quickly found to be incorrect, but that the development of our understanding of atomic structure was not obstructed, which allowed new ideas to build from it. Knowledge is somewhat fickle. If we can accept this and that we will develop our knowledge in the future, we would be less likely to automatically reject new information that conflicts with our current understanding.
Conclusion; What the future of food security could hold for us
“The current agricultural landscape prioritizes quantity rather than quality and chemistry rather than biology… but ending this isn’t impossible” – Peter Behrens
In order to tackle the increasingly large opponent that is endotoxemia and diet-induced chronic disease, the quality of our food, and therefore the food system in its entirety, needs to be confronted directly at the source. It’s not an easy job, but we need to address the complexity head-on.
I believe that regenerative agriculture, primarily through its respect of the reality that healthy soil begets healthy plants and healthy plants beget healthy soil, offers the best foot forward in solving many of our systemic health problems. Regenerative agriculture protects and harnesses soil biota-plant symbiotic relationships (which usually take decades to develop), revoking the need for chemical pesticides and fertilisers, and halting the silent mass-poisoning of people (and plants).
Regenerative farming leverages data to gather insights relating plant diseases to nutrient deficiencies, understanding that pests and pathogens feed only on weak, unhealthy plants. Therefore, plants that are healthy, because their soil contains all the correct components in sufficient quantities, will either require fewer, or completely lack the need for harsh chemical pesticides and synthetic fertilisers.
Many assume regenerative agriculture is not sustainable in the long term because it is not profitable. While easier said than done, this is still far from the truth. A great restaurant and its tasty, high-quality food, attentive service, reasonable prices, and pleasant atmosphere creates happy customers that want to keep coming back for more. Prices don’t need to be insultingly high because the restaurant’s footfall is much higher – and will be for much longer – with this approach. Regenerative farming is much the same. John Kempf articulated this well: “When farmers make wise management decisions based on data, plant health improves, the need for inputs is reduced, and increased yields and profitability necessarily follow”.
In addition to regenerative farming, the importance of small farms and growing your own food cannot be emphasised enough right now. Small family farms are one of our last connections to real food. They’re not inefficient, nor are they outdated. They’re just ‘in the way’. In the way of a food system which is driven by profitable ultra-processed food products that can be mass produced cheaply and stacked high up on a shelf in a warehouse somewhere in Chongqing. If small, local farms disappear, the gap between us the consumer and where food really comes from widens enormously, and the more we depend on the harmful ‘food’ that isn’t really food anymore.
If we could finally taste the fruits of the cutting-edge modern research in ecology, agronomy, physiology, medicine, and nutrition, and offer farmers cost-effective, actionable solutions, we might just be able to set off a global cascade of positive change in food security and public health. By lobbying against ultra-processed foods and other toxins, being open minded to novel ideas about physiology, detaching ourselves from our profit-oriented medical culture, and investing in the provision of healthful environments, the pendulum will swing back in favour of the health of the people, rather than the profit of multinational corporations.
Published version:
References (alphabetical order):
Bánhegyi, G., Mucha, I., Garzó, T., Antoni, F. and Mandl, J. (2000). Endotoxin inhibits glucuronidation in the liver: An effect mediated by intercellular communication. Biochemical Pharmacology, [online] 49(1), pp.65–68. doi:https://doi.org/10.1016/0006-2952(94)00389-4
Bitto, A., Galeano, M., Squadrito, F., Minutoli, L., Polito, F., Dye, J.F., Clayton, E.A., Calò, M., Venuti, F.S., Vaccaro, M. and Altavilla, D. (2008). Polydeoxyribonucleotide improves angiogenesis and wound healing in experimental thermal injury. Critical Care Medicine, 36(5), pp.1594–1602. doi:https://doi.org/10.1097/ccm.0b013e318170ab5c
Camilleri, M. (2019). Leaky gut: mechanisms, measurement and clinical implications in humans. Gut, [online] 68(8), pp.1516–1526. doi:https://doi.org/10.1136/gutjnl-2019-318427
Cao, H. (2023). Bacterial endotoxin lipopolysaccharides regulate gene expression in human colon cancer cells. BMC Research Notes, 16(1). doi:https://doi.org/10.1186/s13104-023-06506-9
Cavalieri, E. and Rogan, E. (2021). The 3,4-Quinones of Estrone and Estradiol Are the Initiators of Cancer whereas Resveratrol and N-acetylcysteine Are the Preventers. International Journal of Molecular Sciences, [online] 22(15), pp.8238–8238. doi:https://doi.org/10.3390/ijms22158238
Christeff, N., Auclair, M.-C., Thobie, N., Benassayag, C. and Nunez, E.A. (2002). Endotoxin induced changes in serum estrogen in male rats: Influence of testicular maturation. Life Sciences, [online] 48(24), pp.2341–2348. doi:https://doi.org/10.1016/0024-3205(91)90271-C
Chou, C.-H., Chen, S.-U., Chen, C.-D., Shun, C.-T., Wen, W.-F., Tu, Y.-A. and Yang, J.-H. (2019). Mitochondrial Dysfunction Induced by High Estradiol Concentrations in Endometrial Epithelial Cells. The Journal of Clinical Endocrinology & Metabolism, [online] 105(1), pp.126–135. doi:https://doi.org/10.1210/clinem/dgz015
Cook, J.A., Wise, W.C., Knapp, D.R. and Halushka, P.V. (1981). Essential fatty acid deficient rats: a new model for evaluating arachidonate metabolism in shock. Advances in Shock Research, [online] 6, pp.93–105. Available at: https://pubmed.ncbi.nlm.nih.gov/6818852/
Do, N.M. (2024). From Leaky Gut to Leaky Skin: A Clinical Review of Lifestyle Influences on the Microbiome. American Journal of Lifestyle Medicine. doi:https://doi.org/10.1177/15598276241292605
Fei, X., Dou, Y., Lv, W., Ding, B., Wei, J., Wu, X., He, X., Fei, Z. and Fei, F. (2022). TLR4 Deletion Improves Cognitive Brain Function and Structure in Aged Mice. Neuroscience, 492, pp.1–17. doi:https://doi.org/10.1016/j.neuroscience.2022.04.007
Flasbeck, V., Dersch, N., Engler, H., Manfred Schedlowski and Brüne, M. (2024). Acute experimental inflammation in healthy women attenuates empathy for psychological pain. Brain, Behavior, and Immunity, 119, pp.1–5. doi:https://doi.org/10.1016/j.bbi.2024.03.032
Ghanim, H., Sia, C.L., Upadhyay, M., Korzeniewski, K., Viswanathan, P., Abuaysheh, S., Mohanty, P. and Dandona, P. (2010). Orange juice neutralizes the proinflammatory effect of a high-fat, high-carbohydrate meal and prevents endotoxin increase and Toll-like receptor expression. The American Journal of Clinical Nutrition, [online] 91(4), pp.940–949. doi:https://doi.org/10.3945/ajcn.2009.28584
Grady, D., Gebretsadik, T., Kerlikowske, K., Ernster, V. and Petitti, D. (1995). Hormone replacement therapy and endometrial cancer risk: a meta-analysis. Obstetrics and Gynecology, [online] 85(2), pp.304–313. doi:https://doi.org/10.1016/0029-7844(94)00383-O
Grigoleit, J.-S., Kullmann, J.S., Wolf, O.T., Hammes, F., Wegner, A., Jablonowski, S., Engler, H., Gizewski, E., Oberbeck, R. and Schedlowski, M. (2011). Dose-Dependent Effects of Endotoxin on Neurobehavioral Functions in Humans. PLoS ONE, [online] 6(12), p.e28330. doi:https://doi.org/10.1371/journal.pone.0028330
Hashida, M., Steelman, A.J. and Erdman, J.W. (2024). α-Tocopherol Depletion Exacerbates Lipopolysaccharide-Induced Reduction of Grip Strength. The Journal of Nutrition, [online] 154(2), pp.498–504. doi:https://doi.org/10.1016/j.tjnut.2023.12.031
Hawiger, J., Hawiger, A., Steckley, S., Timmons, S. and Cheng, C. (1977). Membrane Changes in Human Platelets Induced by Lipopolysaccharide Endotoxin. British Journal of Haematology, 35(2), pp.285–299. doi:https://doi.org/10.1111/j.1365-2141.1977.tb00585.x
Heaney, A.P., Fernando, M. and Melmed, S. (2002). Functional role of estrogen in pituitary tumor pathogenesis. Journal of Clinical Investigation, 109(2), pp.277–283. doi:https://doi.org/10.1172/jci14264
Lafront, C., Germain, L., Campolina-Silva, G.H., Weidmann, C., Berthiaume, L., Hovington, H., Brisson, H., Jobin, C., Frégeau-Proulx, L., Cotau, R., Gonthier, K., Lacouture, A., Caron, P., Ménard, C., Atallah, C., Riopel, J., Latulippe, É., Bergeron, A., Toren, P. and Guillemette, C. (2024). The estrogen signaling pathway reprograms prostate cancer cell metabolism and supports proliferation and disease progression. Journal of Clinical Investigation, 134(11). doi:https://doi.org/10.1172/jci170809
Langdon, S.P., Herrington, C.S., Hollis, R.L. and Gourley, C. (2020). Estrogen Signaling and Its Potential as a Target for Therapy in Ovarian Cancer. Cancers, 12(6), p.1647. doi:https://doi.org/10.3390/cancers12061647
Lin, T., Chang, Y., Wu, M., Wu, T., Chen, C. and Lim, P. (2022). Serum lipopolysaccharide‐binding protein levels and cardiovascular events in hemodialysis patients: A prospective cohort study. Nephrology, 27(11), pp.877–885. doi:https://doi.org/10.1111/nep.14107
Mallardi, D., Maqoud, F., Guido, D., Aloisio, M., Linsalata, M. and Russo, F. (2025). Mapping Research Trends on Intestinal Permeability in Irritable Bowel Syndrome with a Focus on Nutrition: A Bibliometric Analysis. Nutrients, [online] 17(6), pp.1064–1064. doi:https://doi.org/10.3390/nu17061064
Mastrangelo, A., Robles-Vera, I., Mañanes, D., Galán, M., Femenía-Muiña, M., Redondo-Urzainqui, A., Barrero-Rodríguez, R., Papaioannou, E., Amores-Iniesta, J., Devesa, A., Lobo-González, M., Carreras, A., Beck, K.R., Ivarsson, S., Gummesson, A., Georgiopoulos, G., Rodrigo-Tapias, M., Martínez-Cano, S., Fernández-López, I. and Nuñez, V. (2025). Imidazole propionate is a driver and therapeutic target in atherosclerosis. Nature, [online] doi:https://doi.org/10.1038/s41586-025-09263-w
Meléndez-Hevia, E., de Paz-Lugo, P., Cornish-Bowden, A. and Cárdenas, M.L. (2009). A weak link in metabolism: the metabolic capacity for glycine biosynthesis does not satisfy the need for collagen synthesis. Journal of Biosciences, 34(6), pp.853–872. doi:https://doi.org/10.1007/s12038-009-0100-9
Peat, R. (2001). Generative Energy
Perng, W., Friedman, J.E., Janssen, R.C., Glueck, D.H. and Dabelea, D. (2022). Endotoxin Biomarkers Are Associated With Adiposity and Cardiometabolic Risk Across 6 Years of Follow-up in Youth. The Journal of Clinical Endocrinology & Metabolism, [online] 107(7), pp.e3018–e3028. doi:https://doi.org/10.1210/clinem/dgac149
Pogue, A.I., Jaber, V.R., Sharfman, N.M., Zhao, Y. and Lukiw, W.J. (2022). Downregulation of Neurofilament Light Chain Expression in Human Neuronal-Glial Cell Co-Cultures by a Microbiome-Derived Lipopolysaccharide-Induced miRNA-30b-5p. Frontiers in Neurology, [online] 13. doi:https://doi.org/10.3389/fneur.2022.900048
Poto, R., Fusco, W., Rinninella, E., Cintoni, M., Kaitsas, F., Raoul, P., Caruso, C., Mele, M.C., Varricchi, G., Gasbarrini, A., Cammarota, G. and Ianiro, G. (2024). The Role of Gut Microbiota and Leaky Gut in the Pathogenesis of Food Allergy. Nutrients, [online] 16(1), p.92. doi:https://doi.org/10.3390/nu16010092
Radjabzadeh, D., Bosch, J.A., Uitterlinden, A.G., Zwinderman, A.H., Ikram, M.A., van Meurs, J.B.J., Luik, A.I., Nieuwdorp, M., Lok, A., van Duijn, C.M., Kraaij, R. and Amin, N. (2022). Gut microbiome-wide association study of depressive symptoms. Nature Communications, 13(1), p.7128. doi:https://doi.org/10.1038/s41467-022-34502-3
Reifenberg, K., Lehr, H.-A., Fan, J., Koike, T., Wiese, E., Küpper, I., Tolga Atilla Sagban, Schaefer, S.C., Ulrich Zähringer, Torzewski, M., Lackner, K.J. and Sucharit Bhakdi (2009). Endotoxin accelerates atherosclerosis independent of complement activation. Thrombosis Research, 123(4), pp.653–658. doi:https://doi.org/10.1016/j.thromres.2008.06.017
Shan, B., Shao, M., Zhang, Q., Hepler, C., Paschoal, V.A., Barnes, S.D., Lavanya Vishvanath, An, Y.A., Jia, L., Malladi, V.S., Strand, D.W., Gupta, O.T., Elmquist, J.K., Oh, D. and Gupta, R.K. (2020). Perivascular mesenchymal cells control adipose-tissue macrophage accrual in obesity. Nature Metabolism, [online] 2(11), pp.1332–1349. doi:https://doi.org/10.1038/s42255-020-00301-7
Smida, T., Bruno, T.C. and Stabile, L.P. (2020). Influence of Estrogen on the NSCLC Microenvironment: A Comprehensive Picture and Clinical Implications. Frontiers in Oncology, 10. doi:https://doi.org/10.3389/fonc.2020.00137
Steinberg, K.K., Smith, S.J., Thacker, S.B. and Stroup, D.F. (1994). Breast Cancer Risk and Duration of Estrogen Use. Epidemiology, 5(4), pp.415–421. doi:https://doi.org/10.1097/00001648-199407000-00007
Stockmaier, S., Bolnick, D.I., Page, R.A., Josic, D. and Carter, G.G. (2020). Immune-challenged vampire bats produce fewer contact calls. Biology Letters, 16(7), p.20200272. doi:https://doi.org/10.1098/rsbl.2020.0272
Tang, S., Lian, X., Cheng, H., Guo, J., Ni, D., Huang, C., Gu, X., Meng, H., Jiang, J. and Li, X. (2021). Bacterial Lipopolysaccharide Augmented Malignant Transformation and Promoted the Stemness in Prostate Cancer Epithelial Cells. Journal of Inflammation Research, [online] 14, pp.5849–5862. doi:https://doi.org/10.2147/JIR.S332943
Tremellen, K., Syedi, N., Tan, S. and Pearce, K. (2014). Metabolic endotoxaemia – a potential novel link between ovarian inflammation and impaired progesterone production. Gynecological Endocrinology, 31(4), pp.309–312. doi:https://doi.org/10.3109/09513590.2014.994602
Tremellen, K. (2016). Gut Endotoxin Leading to a Decline IN Gonadal function (GELDING) – a novel theory for the development of late onset hypogonadism in obese men. Basic and Clinical Andrology, 26(1). doi:https://doi.org/10.1186/s12610-016-0034-7
Wang, Y.S. and White, T.D. (1999). The Bacterial Endotoxin Lipopolysaccharide Causes Rapid Inappropriate Excitation in Rat Cortex. Journal of Neurochemistry, 72(2), pp.652–660. doi:https://doi.org/10.1046/j.1471-4159.1999.0720652.x
This article has since been published in both The War Kitchen Magazine (Issue 38) and the University of Sheffield’s Biology Magazine.
“If a newly discovered substance had aspirin’s anti-infective, anti-cancer, anti-stress, antioxidant and anti-inflammatory actions, it would be the most researched substance in history.” – Ray Peat, 2013
Aspirin is often viewed through a modern pharmaceutical lens, yet its origins are deeply rooted in ancient plant medicine. Derived from salicylic acid, a natural metabolite from willow bark and a variety of other plants, aspirin has been used for thousands of years to treat inflammation and pain. The Ancient Egyptians used willow leaves for a range of medicinal purposes, and even the Ancient Greek philosopher Hippocrates recommended infusions of willow bark to help reduce fever and ease pain during childbirth. Although Hippocrates wouldn’t have known about salicin (the raw compound in willow, which is metabolised into salicylic acid in the body), this nonetheless serves as a good example of the seemingly lost art of attentive observation. In the modern world of being told everything is either black or white, right or wrong, or this or that, we could all take a page (or quite a few) out of the Ancient Greeks’ book. The late Ray Peat articulated this well: “Perceive, think, act”.
Aspirin and WW1
It wasn’t until 1897 that chemist Felix Hoffmann, working for Bayer in Germany at the time, synthesised acetylsalicylic acid, creating the more stable and less irritating form of the compound we know today as aspirin. France had also been experimenting with salicylic acid derivatives at the time, but it was Hoffman’s addition of an acetyl group which made it a practical and marketable drug, an invention often mistakenly attributed to the French.
Aspirin was revolutionary during the early 20th century, not only because it was more effective and better tolerated than the other over-the-counter pain relievers, but also because it proved to be paramount in treating soldiers’ pain and fevers during WW1. Aspirin was even seen as so effective and important that the patent that Bayer had on aspirin at the time was prematurely broken by the Allied countries during the war. As part of wartime measures, Bayer’s German trademarks and patents were seized in Allied countries and companies in Britain, France, and the USA were allowed to produce and sell aspirin without Bayer’s permission. Aspirin also helped combat the 1918 influenza pandemic, being one of the main remedies widely available, and by the 1920s aspirin was a household staple and firmly established as a standard over-the-counter medicine.
Where did it go wrong?
However, aspirin’s status began to shift in the decades that followed. Aspirin became sold less and less, despite it still being the same-old effective compound. This can be predominantly attributed to the expiration of the patent on the synthesis of aspirin, allowing it to be mass-produced and cheaply made. As a result, aspirin brought in smaller profits, leading to reduced advertising and declining sales. In addition to this, the development of other pain relieving alternatives forced aspirin to slowly fade into obscurity, since pharmaceutical companies had more profitable, competing products to sell.
However, it wasn’t until the 1970s when aspirin had a renaissance. Scientists discovered that aspirin could do more than just kill pain. They found that aspirin could also reduce the risk of heart attacks and strokes, which would prove to be particularly beneficial given the rise in cardiovascular disease at the time (Dalen et al., 2014, Mahmood et al., 2014). A wave of studies followed, and researchers have been making novel discoveries ever since.
Aspirin’s effects
Beyond its common association with pain relief, aspirin’s effects extend to nearly every major biological system – a pharmacological Swiss Army knife. It is known to be anti-inflammatory, anti-cancer, anti-stress, pro-metabolic, neuroprotective, and liver-protective. In terms of hormone regulation and cellular metabolism, it has been shown to lower cortisol, excess lipolysis, aldosterone, excess oestrogen, serotonin, parathyroid hormone, prolactin, and both the formation of prostaglandins and lactic acid.
Aspirin’s most important role, though, may lie in its potent anti-inflammatory properties. It functions by blocking the cyclooxygenase (COX) enzymes, particularly COX-1 and COX-2, thus reducing production of the inflammatory prostaglandins derived from polyunsaturated fatty acids (PUFA). It’s this key mechanism which underpins many of its broader systemic effects.
Aspirin has also demonstrated cognitive-enhancing and mood-boosting effects. It is considered anti-fungal (biofilm-disruptive), an antioxidant, an antihistamine, antiviral, and anabolic. Beyond these, aspirin appears to have several lesser-known but still fascinating effects, such as preventing tooth decay, improving insulin sensitivity, increasing testosterone levels in men, and even extending lifespan.
Aspirin also exhibits a number of paradoxical effects. For example, it is an antioxidant that protects against lipid peroxidation. It can inhibit abnormal cell division, but promote normal cell division (Rothwell et al., 2011). One particularly interesting example of aspirin’s extraordinary effects is how salicylic acid is actually protective to the stomach, intestine, and other organs against the damage caused by other drugs. In the body, acetylsalicylic acid (aspirin) is broken down into acetic acid (found in vinegar) and salicylic acid (found in plants). And, “when aspirin was compared with other common anti-inflammatory drugs, it was found that the salicylic acid it releases protects against the damage done by the other drugs” – Ray Peat, 2006. (Ligumsky et al., 1985; Takeuchi et al., 2001).
For now, and for the future
Aspirin is still one of the most widely studied compounds in the world every year, despite being formulated almost 150 years ago. Even now, we are still uncovering more of aspirin’s remarkable effects. For example, last year research led by Georgi Dinkov showed that, with a combination of aspirin and vitamins B1/B3/B7, the team were able to completely cure 3 of 3 mice from a lethal human tumour (JeKo-1 cell line) with 100% lethality and no reported spontaneous regression. For context, a previous study tested this combination on the same tumour without the aspirin (only the B vitamins) and none of the mice survived, although the increase in tumour volume was perturbed.
And in only March of this year, a study was released showing that aspirin prevents metastasis—when a single cell breaks off from the original tumour into the bloodstream and, like a seed in the wind, spreads to other places in the body (Yang et al., 2025). Fortunately, the body has a mechanism to protect itself against metastasis in the form of T-cells. T-cells are able to recognise and destroy these cancerous cells. However, platelets that typically stop bleeding can suppress T-cells and prevent them from destroying these cancerous cells. Aspirin was observed to disrupt these platelets and allow the T-cells to “hunt down” the cancer cells, unleashing the T-cells onto the cancer.
Furthermore, a groundbreaking study was recently released, assessing the long term impacts of low dose aspirin on cancer prevention. It reviewed the cancer incidence from a whopping 1,506,525 people living in Hong Kong across 20 years, with 538,147 taking 80mg of aspirin per day (a very small dose), and the rest being the control group. The control group cohorts were ranked as being much healthier across the board than the aspirin group (15x fewer strokes, 12x fewer incidences of coronary heart disease, twice as less cases of hypertension etc). Over 10 years, aspirin users only had a slightly smaller prevalence of cancer (6.9% vs. 7.1%, however, those taking 80mg of aspirin for longer than 10 years had a significantly lower cancer risk (4.6% lower, p < 0.05). And remember, this is a 500,000-large group of people significantly less healthy than the control group. Aspirin users also had fewer deaths in total, fewer deaths from cancers, and a reduced risk of every cancer, besides bladder and leukemia. If you account for the poor health the aspirin group was already in at the start of the study, it cut cancer risk by around 50%, an enormous figure.
Conclusion
Far from being a relic of early 20th-century medicine, aspirin continues to prove itself as one of the most versatile and powerful therapeutic agents discovered to date. Its transformation from an ancient herbal remedy to a marvel of modern medicine reflects not only its effectiveness at a biochemical level but also its evolutionary alignment with human physiology; the many “unexpected” beneficial effects of aspirin strongly suggest that it is acting on fundamental biological processes in a positive and coherent way. Aspirin blurs the line between drug and nutrient, synthetic and natural, and perhaps we’re only just beginning to understand the true scope of its potential.
There hasn’t been a year since 2005, where fewer than 2,000 research papers have been published on aspirin, and the number is steadily growing as each year passes by. There seems to be a positive trend in this field of research, and partly from it emerges a collective pursuit of truly understanding the complexity of human physiology, led by the people, for the health of the people. A sizable proportion of the “Ray Peat sphere” on ‘X’ has been, and still is, a breeding ground for ideas and discussion about nutrition and bioenergetics, and is helping to accelerate this pursuit. As research continues to unveil aspirin’s effects, it challenges us to reconsider whether the most effective therapies might still lie in mother nature and the wisdom of the past.
References:
General:
Takeuchi, K., Hase, S., Mizoguchi, H., Yusaku Komoike and Tanaka, A. (2001). Protection by aspirin of indomethacin-induced small intestinal damage in rats: mediation by salicylic acid. Journal of Physiology-Paris, 95(1-6), pp.51–57.
M. Ligumsky, Guth, P.H., J. Elashoff, Kauffman, G.L., Hansen, D. and Paulsen, G. (1985). Salicylic Acid Blocks Indomethacin-Induced Cyclooxygenase Inhibition and Lesion Formation in Rat Gastric Mucosa. Experimental Biology and Medicine, 178(2), pp.250–253. doi:https://doi.org/10.3181/00379727-178-42007.
Mahmood, S.S., Levy, D., Vasan, R.S. and Wang, T.J. (2014). The Framingham Heart Study and the epidemiology of cardiovascular disease: a historical perspective. The Lancet, 383(9921), pp.999–1008. doi:https://doi.org/10.1016/s0140-6736(13)61752-3.
Dalen, J.E., Alpert, J.S., Goldberg, R.J. and Weinstein, R.S. (2014). The Epidemic of the 20th Century: Coronary Heart Disease. The American Journal of Medicine, [online] 127(9), pp.807–812. doi:https://doi.org/10.1016/j.amjmed.2014.04.015.
Anti-cancer:
Rothwell, P.M., Wilson, M., Price, J.F., Belch, J.F., Meade, T.W. and Mehta, Z. (2012). Effect of daily aspirin on risk of cancer metastasis: a study of incident cancers during randomised controlled trials. The Lancet, [online] 379(9826), pp.1591–1601. doi:https://doi.org/10.1016/s0140-6736(12)60209-8.
Mills, E.J., Wu, P., Alberton, M., Kanters, S., Lanas, A. and Lester, R. (2012). Low-dose Aspirin and Cancer Mortality: A Meta-analysis of Randomized Trials. The American Journal of Medicine, 125(6), pp.560–567. doi:https://doi.org/10.1016/j.amjmed.2012.01.017.
Arango, H.A., Icely, S., Roberts, W.S., Cavanagh, D. and Becker, J.L. (2001). Aspirin effects on endometrial cancer cell growth. Obstetrics and Gynecology, [online] 97(3), pp.423–427. doi:https://doi.org/10.1016/s0029-7844(00)01161-3.
Elwood, P.C., Morgan, G., Delon, C., Protty, M., Galante, J., Pickering, J., Watkins, J., Weightman, A. and Morris, D. (2021). Aspirin and cancer survival: a systematic review and meta-analyses of 118 observational studies of aspirin and 18 cancers. ecancermedicalscience, 15. doi:https://doi.org/10.3332/ecancer.2021.1258.
Lin, Q., Bai, M.-J., Wang, H.-F., Wu, X.-Y., Huang, M.-S. and Li, X. (2021). Aspirin-induced long-term tumor remission in hepatocellular carcinoma with adenomatous polyposis coli stop-gain mutation: A case report. World Journal of Clinical Cases, 9(24), pp.7189–7195. doi:https://doi.org/10.12998/wjcc.v9.i24.7189.
Wang, L., Zhang, R., Yu, L., Xiao, J., Zhou, X., Li, X., Song, P. and Li, X. (2021). Aspirin Use and Common Cancer Risk: A Meta-Analysis of Cohort Studies and Randomized Controlled Trials. Frontiers in Oncology, 11. doi:https://doi.org/10.3389/fonc.2021.690219.
Jiang, M.-C., Liao, C.-F. and Lee, P.-H. (2001). Aspirin Inhibits Matrix Metalloproteinase-2 Activity, Increases E-Cadherin Production, and Inhibits in Vitro Invasion of Tumor Cells. Biochemical and Biophysical Research Communications, 282(3), pp.671–677. doi:https://doi.org/10.1006/bbrc.2001.4637.
Yang, J., Yamashita-Kanemaru, Y., Morris, B.I., Contursi, A., Trajkovski, D., Xu, J., Patrascan, I., Benson, J., Evans, A.C., Conti, A.G., Al-Deka, A., Dahmani, L., Avdic-Belltheus, A., Zhang, B., Okkenhaug, H., Whiteside, S.K., Imianowski, C.J., Wesolowski, A.J., Webb, L.V. and Puccio, S. (2025). Aspirin prevents metastasis by limiting platelet TXA2 suppression of T cell immunity. Nature, [online] pp.1–10.
Lowers cortisol:
Nixon, M., Wake, D.J., Livingstone, D.E., Stimson, R.H., Esteves, C.L., Seckl, J.R., Chapman, K.E., Andrew, R. and Walker, B.R. (2012). Salicylate Downregulates 11β-HSD1 Expression in Adipose Tissue in Obese Mice and in Humans, Mediating Insulin Sensitization. Diabetes, 61(4), pp.790–796. doi:https://doi.org/10.2337/db11-0931.
Rangasamy, S.B., Dasarathi, S., Pahan, P., Jana, M. and Pahan, K. (2019). Low-dose aspirin upregulates tyrosine hydroxylase and increases dopamine production in dopaminergic neurons: Implications for Parkinson’s disease. Journal of neuroimmune pharmacology : the official journal of the Society on NeuroImmune Pharmacology, [online] 14(2), pp.173–187. doi:https://doi.org/10.1007/s11481-018-9808-3.
Lowers serotonin:
Vitale, G., Pini, L.-A., Ottani, A. and Maurizio Sandrini (1998). Effect of Acetylsalicylic Acid on Formalin Test and on Serotonin System in the Rat Brain. General Pharmacology The Vascular System, 31(5), pp.753–758. doi:https://doi.org/10.1016/s0306-3623(98)00108-6.
Sandrini, M., Vitale, G., Dondi, M. and Pini, L.A. (1995). Effects of acetylsalicylic acid on serotonin brain receptor subtypes. General Pharmacology: The Vascular System, 26(4), pp.737–741. doi:https://doi.org/10.1016/0306-3623(94)00252-i.
Lowers prolactin:
Luigi Di Luigi, Guidetti, L., Romanelli, F., Baldari, C. and Conte, D. (2001). Acetylsalicylic acid inhibits the pituitary response to exercise-related stress in humans. Medicine and Science in Sports and Exercise, 33(12), pp.2029–2035. doi:https://doi.org/10.1097/00005768-200112000-00009.
Lowers excess lipolysis:
Erman, A., Schwartzman, M. and Raz, A. (1980). Indomethacin but not aspirin inhibits basal and stimulated lipolysis in rabbit kidney. Prostaglandins, [online] 20(4), pp.689–702. doi:https://doi.org/10.1016/0090-6980(80)90108-2.
Lowers parathyroid hormone:
Alkhalaf, Z., Kim, K., Kuhr, D.L., Radoc, J.G., Purdue-Smithe, A., Pollack, A.Z., Yisahak, S.F., Silver, R.M., Thoma, M., Kissell, K., Perkins, N.J., Sjaarda, L.A. and Mumford, S.L. (2021). Markers of vitamin D metabolism and premenstrual symptoms in healthy women with regular cycles. Human Reproduction, 36(7), pp.1808–1820. doi:https://doi.org/10.1093/humrep/deab089.
Lowers aldosterone:
Snoep, J.D., Hovens, M.M.C., Pasha, S.M., Frölich, M., Pijl, H., Tamsma, J.T. and Huisman, M.V. (2009). Time-dependent effects of low-dose aspirin on plasma renin activity, aldosterone, cortisol, and catecholamines. Hypertension (Dallas, Tex. : 1979), [online] 54(5), pp.1136–42. doi:https://doi.org/10.1161/HYPERTENSIONAHA.109.134825.
Lowers prostaglandin formation:
Barnes, C.J., Hamby-Mason, R.L., Hardman, W.E., Cameron, I.L., Speeg, K.V. and Lee, M. (1999). Effect of aspirin on prostaglandin E2 formation and transforming growth factor alpha expression in human rectal mucosa from individuals with a history of adenomatous polyps of the colon. Cancer epidemiology, biomarkers & prevention : a publication of the American Association for Cancer Research, cosponsored by the American Society of Preventive Oncology, [online] 8(4 Pt 1), pp.311–5. Available at: https://pubmed.ncbi.nlm.nih.gov/10207634/.
Lowers lactic acid formation:
Ren, G., Ma, Y., Wang, X., Zheng, Z. and Li, G. (2022). Aspirin blocks AMPK/SIRT3-mediated glycolysis to inhibit NSCLC cell proliferation. European Journal of Pharmacology, [online] p.175208. doi:https://doi.org/10.1016/j.ejphar.2022.175208.
Pro-metabolic:
Mahmud, T., Rafi, S.S., Scott, D.L., Wrigglesworth, J.M. and Bjarnason, I. (1996). Nonsteroidal antiinflammatory drugs and uncoupling of mitochondrial oxidative phosphorylation. Arthritis & Rheumatism, 39(12), pp.1998–2003. doi:https://doi.org/10.1002/art.1780391208.
Meex, R.C.R., Phielix, E., Moonen-Kornips, E., Schrauwen, P. and Hesselink, M.K.C. (2011). Stimulation of human whole-body energy expenditure by salsalate is fueled by higher lipid oxidation under fasting conditions and by higher oxidative glucose disposal under insulin-stimulated conditions. The Journal of clinical endocrinology and metabolism, [online] 96(5), pp.1415–23. doi:https://doi.org/10.1210/jc.2010-1816.
Nootropic:
Ghosh, A., Dhumal, V.R., Tilak, A.V., Das, N., Singh, A. and Bondekar, A.A. (2011). Evaluation of nootropic and neuroprotective effects of low dose aspirin in rats. Journal of Pharmacology and Pharmacotherapeutics, 2(1), pp.3–6. doi:https://doi.org/10.4103/0976-500x.77079.
Mood booster:
Dominiak, M., Gędek, A., Sikorska, M., Mierzejewski, P., Wojnar, M. and Antosik-Wójcińska, A.Z. (2022). Acetylsalicylic Acid and Mood Disorders: A Systematic Review. Pharmaceuticals, 16(1), p.67. doi:https://doi.org/10.3390/ph16010067.
Neuroprotective:
Moro, M.A., J De Alba, A Cárdenas, J De Cristóbal, Leza, J.C., I Lizasoain, M.J.M Dı́az-Guerra, L Boscá and Lorenzo, P. (2000). Mechanisms of the neuroprotective effect of aspirin after oxygen and glucose deprivation in rat forebrain slices. Neuropharmacology, 39(7), pp.1309–1318. doi:https://doi.org/10.1016/s0028-3908(99)00226-9.
Gomes, I. (1998). Aspirin: a neuroprotective agent at high doses? The National medical journal of India, [online] 11(1), pp.14–7. Available at: https://pubmed.ncbi.nlm.nih.gov/9557513/.
Asanuma, M., Nishibayashi-Asanuma, S., Miyazaki, I., Kohno, M. and Ogawa, N. (2001). Neuroprotective effects of non-steroidal anti-inflammatory drugs by direct scavenging of nitric oxide radicals. Journal of Neurochemistry, 76(6), pp.1895–1904. doi:https://doi.org/10.1046/j.1471-4159.2001.00205.x.
Pro-dopamine:
Aubin, N., Curet, O., Deffois, A. and Carter, C. (1998). Aspirin and salicylate protect against MPTP-induced dopamine depletion in mice. Journal of Neurochemistry, [online] 71(4), pp.1635–1642. doi:https://doi.org/10.1046/j.1471-4159.1998.71041635.x.
Liver protective:
F. Edward Boas, Brown, K.T., Ziv, E., Hooman Yarmohammadi, Sofocleous, C.T., Erinjeri, J.P., Harding, J.J. and Solomon, S.B. (2019). Aspirin Is Associated With Improved Liver Function After Embolization of Hepatocellular Carcinoma. American Journal of Roentgenology, [online] 213(3), pp.1–7. doi:https://doi.org/10.2214/ajr.18.20846.
Han, Y.-M., Lee, Y.-J., Jang, Y.-N., Kim, H.-M., Seo, H.S., Jung, T.W. and Jeong, J.H. (2020). Aspirin Improves Nonalcoholic Fatty Liver Disease and Atherosclerosis through Regulation of the PPARδ-AMPK-PGC-1α Pathway in Dyslipidemic Conditions. BioMed Research International, [online] 2020, p.7806860. doi:https://doi.org/10.1155/2020/7806860.
Simon, T.G., Henson, J., Osganian, S., Masia, R., Chan, A.T., Chung, R.T. and Corey, K.E. (2019). Daily aspirin use associated with reduced risk for fibrosis progression in patients with nonalcoholic fatty liver disease. Clinical gastroenterology and hepatology : the official clinical practice journal of the American Gastroenterological Association, [online] 17(13), pp.2776-2784.e4. doi:https://doi.org/10.1016/j.cgh.2019.04.061.
Anti-fungal (biofilm disruptive):
Trofa, D., Agovino, M., Stehr, F., Schäfer, W., Rykunov, D., Fiser, A., Hamari, Z., Nosanchuk, J.D. and Gácser, A. (2009). Acetylsalicylic acid (aspirin) reduces damage to reconstituted human tissues infected with Candida species by inhibiting extracellular fungal lipases. Microbes and Infection, [online] 11(14-15), pp.1131–1139. doi:https://doi.org/10.1016/j.micinf.2009.08.007.
Liu, X., Wang, D., Yu, C., Li, T., Liu, J. and Sun, S. (2016). Potential Antifungal Targets against a Candida Biofilm Based on an Enzyme in the Arachidonic Acid Cascade—A Review. Frontiers in Microbiology, 7. doi:https://doi.org/10.3389/fmicb.2016.01925.
Alem, M.A.S. and Douglas, L.J. (2004). Effects of Aspirin and Other Nonsteroidal Anti-Inflammatory Drugs on Biofilms and Planktonic Cells of Candida albicans. Antimicrobial Agents and Chemotherapy, 48(1), pp.41–47. doi:https://doi.org/10.1128/aac.48.1.41-47.2004.
Ellero-Simatos, S., Beitelshees, A.L., Lewis, J.P., Yerges-Armstrong, L.M., Georgiades, A., Dane, A., Harms, A.C., Strassburg, K., Guled, F., Hendriks, M.M.W.B., Horenstein, R.B., Shuldiner, A.R., Hankemeier, T., Kaddurah-Daouk, R. and Pharmacometabolomics Research Network (2015). Oxylipid Profile of Low-Dose Aspirin Exposure: A Pharmacometabolomics Study. Journal of the American Heart Association, [online] 4(10), p.e002203. doi:https://doi.org/10.1161/JAHA.115.002203.
Anabolic:
Chin, K.-Y. (2017). A review on the relationship between aspirin and bone health. Journal of Osteoporosis, [online] 2017, pp.1–8. doi:https://doi.org/10.1155/2017/3710959.
Antiviral:
Glatthaar-Saalmüller, B., Mair, K.H. and Saalmüller, A. (2016). Antiviral activity of aspirin against RNA viruses of the respiratory tract-an in vitro study. Influenza and Other Respiratory Viruses, 11(1), pp.85–92. doi:https://doi.org/10.1111/irv.12421.
Paradoxical effects:
Rothwell, P.M., Fowkes, F.G.R., Belch, J.F., Ogawa, H., Warlow, C.P. and Meade, T.W. (2011). Effect of daily aspirin on long-term risk of death due to cancer: analysis of individual patient data from randomised trials. The Lancet, 377(9759), pp.31–41. doi:https://doi.org/10.1016/s0140-6736(10)62110-1.
Tewari, S., Kaushish, R., Sharma, S. and Gulati, N. (1997). Role of low dose aspirin in prevention of pregnancy induced hypertension. Journal of the Indian Medical Association, [online] 95(2), pp.43–4, 47. Available at: https://pubmed.ncbi.nlm.nih.gov/9357241/.
Tooth decay prevention:
Yuan, M., Zhan, Y., Hu, W., Li, Y., Xie, X., Miao, N., Jin, H. and Zhang, B. (2018). Aspirin promotes osteogenic differentiation of human dental pulp stem cells. International Journal of Molecular Medicine. doi:https://doi.org/10.3892/ijmm.2018.3801.
Stroke and DVT prevention:
Schrör, K. (1997). Aspirin and Platelets: The Antiplatelet Action of Aspirin and Its Role in Thrombosis Treatment and Prophylaxis. Seminars in Thrombosis and Hemostasis, [online] 23(04), pp.349–356. doi:https://doi.org/10.1055/s-2007-996108.
Lavu, M.S., Porto, J.R., Hecht, C.J., Acuña, A.J., Kaelber, D.C., Parvizi, J. and Kamath, A.F. (2024). Low-Dose Aspirin Is the Safest Prophylaxis for Prevention of Venous Thromboembolism After Total Knee Arthroplasty Across All Patient Risk Profiles. The Journal of bone and joint surgery. American volume, [online] 106(14), pp.1256–1267. doi:https://doi.org/10.2106/JBJS.23.01158.
Improving insulin sensitivity:
Yuan, M. (2001). Reversal of Obesity- and Diet-Induced Insulin Resistance with Salicylates or Targeted Disruption of Ikkbeta. Science, 293(5535), pp.1673–1677. doi:https://doi.org/10.1126/science.1061620.
Increasing testosterone levels in men:
L. Di Luigi, Rossi, C., P. Sgrò, Fierro, V., Romanelli, F., C. Baldari and Guidetti, L. (2007). Do Non-Steroidal Anti-Inflammatory Drugs Influence the Steroid Hormone Milieu in Male Athletes? International Journal of Sports Medicine, 28(10), pp.809–814. doi:https://doi.org/10.1055/s-2007-964991.
Extending lifespan:
Berkel, C. and Cacan, E. (2021). A collective analysis of lifespan-extending compounds in diverse model organisms, and of species whose lifespan can be extended the most by the application of compounds. Biogerontology, 22(6), pp.639–653. doi:https://doi.org/10.1007/s10522-021-09941-y.